

文献题目:Salt-stress-induced tomato sweetening involves an SlSnRK2.6-SlZHD8 sugar accumulation cascade triggered by root-derived abscisic acid
发表期刊:THE EMBO JOURNAL
影响因子:8.3
物种:番茄
发表时间:2026年2月2日
发表单位:山东农业大学
蓝景科信提供:DAP-seq技术服务
主要研究结果
“顺境出产量,逆境促品质",这句古老农谚在番茄种植中早已被验证——适度盐碱环境种出的番茄,甜度更高、风味更浓。但背后的科学机制,长期以来是未解之谜。
为解析盐胁迫提升番茄果实糖含量的分子机制,本研究通过同位素示踪与嫁接实验证实,根系来源的ABA是介导盐胁迫诱导果实糖积累的核心信号。进一步基因功能验证表明,SlSnRK2.6激酶作为ABA通路下游关键组分,直接调控盐胁迫下的果实糖积累。酵母双杂交实验确认,SlZHD8是SlSnRK2.6的直接互作蛋白,且对盐胁迫诱导的糖积累起负调控作用。在此基础上,磷酸化实验证明,盐胁迫下根系来源的ABA可促进SlSnRK2.6对SlZHD8的磷酸化修饰,该修饰显著降低SlZHD8的蛋白稳定性;遗传与表型分析进一步证实,完整的SlSnRK2.6-SlZHD8信号模块是盐胁迫下番茄果实糖积累的必需调控单元。
研究通过DAP-seq鉴定SlZHD8在全基因组范围内结合位点,两个重复共获得95,682个可信重叠峰,其中16.9%的峰位于启动子区,且在TSS上游约300bp的近端启动子区高度富集,其核心结合基序为TAATTAAT。GO与KEGG富集分析显示,这些靶基因显著富集于淀粉与蔗糖代谢、蔗糖响应、水解酶活性及液泡膜定位等糖代谢相关过程。通过DAP-seq与RNA-seq联合分析,筛选出SlSUS3(蔗糖水解)和SlSWEET12(糖转运)为SlZHD8调控果实糖积累的关键靶基因。后续经EMSA、ChIP-qPCR、双荧光素酶实验证实,SlZHD8可直接结合二者启动子并发挥转录抑制作用,而SlSnRK2.6在T247位点磷酸化SlZHD8可解除该抑制,进而促进果实糖积累。此外,SlSnRK2.6-SlZHD8-SlSWEET12模块可同步调控根系糖分分配与Na+/K+稳态,参与番茄植株耐盐性调控。群体遗传分析表明,ZHD8在番茄驯化过程中受到人工选择,并从中鉴定出能同时提升果实糖含量与耐盐性的优异天然单倍型。
本研究系统阐明了盐胁迫下根系信号调控果实增甜的分子机制,为逆境促品质这一重要农业现象提供了核心理论支撑,并为番茄品质与抗逆协同改良提供了关键基因资源与育种靶点。
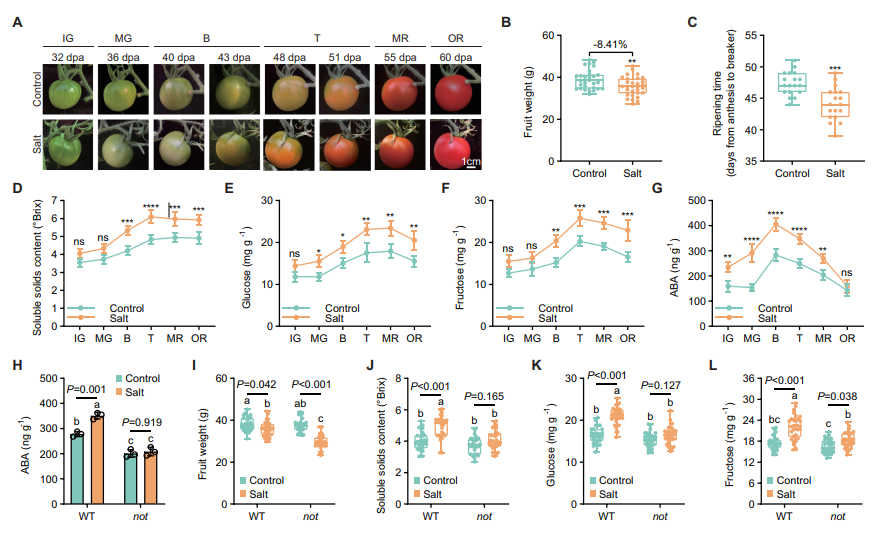
图1. ABA含量升高介导盐胁迫下番茄果实的糖分积累。(A–G)盐胁迫对(A)果实表型、(B)单果重、(C)成熟时间、(D)可溶性固形物含量、(E)葡萄糖含量、(F)果糖含量及(G)ABA含量的影响。(H–L)内源ABA与盐胁迫的互作对(H)ABA含量、(I)单果重、(J)可溶性固形物含量、(K)葡萄糖含量及(L)果糖含量的影响。

图2. 根系来源的ABA激活SlSnRK2.6激酶,介导盐胁迫下番茄果实的糖分积累。(A)盐胁迫对ABA从根/叶向果实运输的影响。(B)对照和盐胁迫条件下,野生型嫁接植株(WT/WT)与未嫁接植株(WT/not)的ABA含量。(C–F)根系来源ABA与盐胁迫的互作对(C)可溶性固形物含量、(D)葡萄糖含量、(E)果糖含量及(F)SlSnRK2.6激酶活性的影响。(G–J)SlSnRK2.6与盐胁迫的互作对(G)单果重、(H)可溶性固形物含量、(I)葡萄糖含量及(J)果糖含量的影响。

图3. SlSnRK2.6与SlZHD8相互作用并使其磷酸化,进而介导盐胁迫下番茄果实的糖分积累。(A)SlZHD8的亚细胞定位。(B)酵母双杂交(Y2H)实验表明SlSnRK2.6与SlZHD8相互作用。(C)Pull-down实验表明SlSnRK2.6与SlZHD8相互作用。(D)免疫共沉淀(Co-IP)实验表明SlSnRK2.6与SlZHD8相互作用。(E)双分子荧光互补(BiFC)实验表明SlSnRK2.6与SlZHD8相互作用。(F)荧光素酶互补成像(LCI)实验表明SlSnRK2.6与SlZHD8相互作用。(G)SlSnRK2.6在体外磷酸化SlZHD8的T247位点。(H)SlSnRK2.6在体内磷酸化SlZHD8的T247位点。(I)盐胁迫下根系来源的ABA参与SlZHD8的体内磷酸化。(J)盐胁迫下SlSnRK2.6参与SlZHD8的体内磷酸化。(K)无细胞降解实验表明SlZHD8发生依赖于SlSnRK2.6的降解。(L)体内实验表明SlZHD8发生依赖于SlSnRK2.6的降解。(M–P)SlSnRK2.6-SlZHD8模块对盐胁迫下果实糖分积累至关重要。(M)单果重、(N)可溶性固形物含量、(O)葡萄糖含量、(P)果糖含量。

图4. SlSnRK2.6介导的SlZHD8磷酸化解除其对SlSUS3和SlSWEET12的转录抑制。(A)韦恩图展示DAP-seq鉴定得到的结合峰。(B)SlZHD8结合峰在番茄基因组上的分布。(C)启动子区域富集的SlZHD8结合基序。(D)启动子区域SlZHD8结合峰的KEGG富集分析。(E)韦恩图展示RNA-seq与DAP-seq的重叠基因情况。(F、G)对照与盐胁迫条件下,SlSUS3和在野生型及slzhd8突变体果实中的表达模式。(H)SlSUS3和SlSWEET12启动子上的SlZHD8结合基序。(I)EMSA实验证实SlZHD8可直接结合SlSUS3和SlSWEET12启动子探针。(J)EMSA实验显示,SlSnRK2.6对SlZHD8的磷酸化降低其DNA结合能力。(K)ChIP实验显示,SlSnRK2.6对SlZHD8的磷酸化降低其DNA结合能力。(L)转录激活实验显示,SlSnRK2.6对SlZHD8的磷酸化解除其转录抑制作用。(M)SlSnRK2.6对SlZHD8的磷酸化解除其对SlSUS3和SlSWEET12的转录抑制。

图5. SlZHD8-SlSUS3/SlSWEET12模块参与盐胁迫下番茄果实糖分积累与胁迫耐受性。(A–C)SlZHD8-SlSUS3模块对盐胁迫下果实糖分积累至关重要。(A)可溶性固形物含量;(B)葡萄糖含量;(C)果糖含量。(D–F)SlZHD8-SlSWEET12模块对盐胁迫下果实糖分积累至关重要。(D)可溶性固形物含量;(E)葡萄糖含量;(F)果糖含量。(G–K)SlZHD8-SlSWEET12模块参与盐胁迫耐受性调控。(G)盐胁迫下幼苗及根系表型;(H)植株干重;(I)根冠比;(J)根系可溶性糖含量;(K)根系Na+/K+比值。

图6. ZHD8的自然变异与果实糖分积累及耐盐性相关。(A)ZHD8位于番茄驯化选择区间内,但未达到改良选择区间的阈值。(B)基于631份番茄种质(包括野生醋栗番茄SP、樱桃番茄SLC、栽培番茄SLL,含传统品种与现代品种)的ZHD8单倍型聚类分析。(C)EMSA实验显示ZHD8HapB单倍型DNA结合能力降低。(D)ChIP实验显示ZHD8HapB单倍型DNA结合能力降低(E)ZHD8单倍型在SP、SLC、SLL(传统品种与现代品种)群体中的频率分布,标注各组中HapA与HapB的数量。(F)ZHD8HapA与ZHD8HapB的地理分布。(G–J)ZHD8HapB单倍型在盐胁迫下增强果实糖分积累。(G)单果重;(H)可溶性固形物含量;(I)葡萄糖含量;(J)果糖含量。(K–O)ZHD8HapB单倍型增强植株耐盐性。(K)盐胁迫下幼苗及根系表型;(L)植株干重;(M)根冠比;(N)根系可溶性糖含量;(O)根系Na+/K+比值。

图7. 机制模式图。在盐胁迫条件下,根部产生的脱落酸(ABA)向果实的运输量增加。果实中升高的ABA会激活SlSnRK2.6激酶,该激酶通过磷酸化SlZHD8并降低其蛋白质稳定性及DNA结合能力,从而抑制SlZHD8的功能;这一过程解除了SlZHD8对SlSUS3和SlSWEET12的抑制作用,进而促进果实糖分积累。与此同时,SlSnRK2.6-SlZHD8-SlSWEET12调控模块还参与调节根部糖分积累并赋予植物盐胁迫耐受性。进化分析鉴定出一种优异单倍型ZHD8HapB,因其启动子结合亲和力降低,可在提升果实糖分的同时增强耐盐性。

